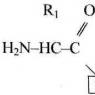
Особенности строения и функции гипоталамуса. Гипоталамус что это такое и за что он отвечает. За что отвечает
ГИПОТАЛАМУС [hypothalamus (BNA, JNA, PNA); греч, hypo- + thalamos комната; син.: подбугорье, гипоталамическая область, подбугровая область ] - отдел промежуточного мозга, расположенный книзу от таламуса под гипоталамической бороздой и представляющий собой скопление нервных клеток с многочисленными афферентными и эфферентными связями.
История
Начиная с середины 19 в. изучалось влияние Г. на различные стороны жизнедеятельности организма (процессы адаптации, половые функции, процессы обмена, теплорегуляции, водно-солевой обмен и др.).
Большой вклад в изучение Г. внесли отечественные ученые. В 30-х годах 20 в. А. Д. Сперанский с сотр. проводил опыты на животных, накладывая стеклянную бусинку или металлическое кольцо на вещество мозга в области турецкого седла, в результате возникали кровоизлияния и язвы в желудке и кишечнике.
H. Н. Бурденко и Б. Н. Могильницкий описали возникновение прободной язвы желудка при нейрохирургическом вмешательстве в области III желудочка. Особое место занимают исследования, проведенные Н. И. Гращенковым при изучении теоретических и клин, аспектов роли Г. при различных расстройствах нервной системы и внутренних органов.
В 1912 г. Ашнер (В. Aschner) наблюдал атрофию гонад у собак после разрушения Г. В 1928 г. Шаррер (В. Scharrer) обнаружил секреторную активность гипоталамических ядер. Хольвег и Юнкман (W. Hohlweg, К. Junkman, 1932) установили локализацию в Г. полового центра, электрическая стимуляция к-рого в опытах Харриса (G. W. Harris, 1937) вызывала овуляцию у кроликов. В 1950 г. Хьюм и Виттенштейн (D. М. Hume, G. J. Wittenstein) показали влияние гипоталамических экстрактов на секрецию адренокортикотропного гормона. В 1955 г. Гиллемин и Розенберг (R. Guillemin, В. Rosenberg) обнаружили в Г. так наз. освобождающий фактор - кортикотропин (кортикотропин-рилизинг-фактор). В последующие годы была показана локализация ядер некоторых Г., ответственных за регуляцию обмена веществ и секрецию отдельных гормонов гипофиза (см.).
Эмбриология, анатомия, гистология
Г. является филогенетически древним образованием, существующим у всех хордовых. Однако обозначение этого отдела головного мозга как подбугорье не может быть использовано в отношении круглоротых и поперечноротых, т. к. зрительные бугры впервые формируются на стадии амфибий. У птиц Г. имеет относительно небольшой размер, но дифференциация его ядер выражена достаточно хорошо. Он получает в основном импульсы из обонятельных центров, полосатого тела, образующего у птиц большую часть переднего мозга.
Наивысшего развития Г. достигает у млекопитающих. У эмбриона человека в возрасте 3 мес. на внутренней поверхности таламуса имеются две борозды, разделяющие его на три части: верхнюю - эпиталамус, среднюю - таламус и нижнюю - гипоталамус. В дальнейшем эмбриональном развитии выявляется более тонкая дифференциация ядер Г. и формируются его многочисленные связи. Передней границей Г. является зрительный перекрест (chiasma opticum) , терминальная пластинка (lamina terminalis) и передняя спайка (commissura ant.). Задняя граница проходит позади нижнего края сосцевидных тел (corpora mamillaria). Кпереди клеточные группы Г. без перерыва переходят в клеточные группы пластинки прозрачной перегородки (lamina septi pellucidi). Несмотря на небольшие размеры Г., его цитоархитектоника отличается значительной сложностью. В Г. хорошо развито серое вещество, состоящее гл. обр. из мелких клеток. В некоторых участках лежат группы клеток, образующие отдельные ядра Г. (рис. 1). Число, топография, размер, форма и степень дифференцировки этих ядер варьируют у разных позвоночных; у млекопитающих обычно различают 32 пары ядер. Между соседними ядрами существуют промежуточные нервные клетки или их небольшие группы, поэтому физиол. значение могут иметь не только ядра, но и некоторые межъядерные Гипоталамические зоны. По группировке в Г. условно различают три нерезко разграниченные области скопления ядер: переднюю, среднюю и заднюю.
В средней области Г., вокруг нижнего края III желудочка, лежат серобугорные ядра (nucll. tuberales), дуговидно охватывающие воронку (infundibulum). Кверху и немного латеральнее от них лежат крупные верхне-медиальные и нижне-медиальные ядра. Нервные клетки, составляющие эти ядра, неодинаковы по размеру. На периферии локализуются мелкие нервные клетки, а более крупные занимают середину ядер. Нервные клетки верхне-медиального и нижне-медиального ядер отличаются друг от друга строением дендритов. У клеток верхне-медиальных ядер дендриты характеризуются наличием большого числа длинных шипиков, аксоны сильно разветвляются и имеют многочисленные синаптические связи. Серобугорные ядра (nucll. tuberales) представляют собой скопления небольших нервных клеток веретеновидной или треугольной формы, локализующиеся вокруг основания воронки. Отростки нервных клеток этих ядер определяются в проксимальной части гипофизарной ножки до срединного возвышения, где они заканчиваются аксовазальными синапсами на петлях первичной капиллярной сети гипофиза. Эти клетки дают начало волокнам туберогипофизарного пучка.
Группа ядер задней области состоит из рассеянных крупных клеток, среди которых лежат скопления мелких клеток. К этому отделу относятся также ядра сосцевидного тела (nucll. corporis mamillaris), которые выступают на нижней поверхности промежуточного мозга в виде полушарий (парных у приматов и непарных у прочих млекопитающих). Клетки этих ядер являются эфферентными нервными клетками и дают начало одной. из главнейших проекционных систем из Г. в продолговатый и спинной мозг. Наиболее крупное клеточное скопление образует медиальное ядро сосцевидного тела. Кпереди от сосцевидных тел выступает дно III желудочка в виде серого бугра (tuber cinereum), образованного тонкой пластинкой серого вещества. Этот выступ вытягивается в воронку, переходящую в дисталь-пом направлении в гипофизарную ножку и далее в заднюю долю гипофиза. Воронка отграничивается от серого бугра неясно выраженной бороздой. Расширенная верхняя часть воронки - срединное возвышение- имеет особое строение и своеобразную васкуляризации). Со стороны полости воронки срединное возвышение выстлано эпендимой, за к-рой идет слой нервных волокон гипоталамо-гипофизарного пучка и более тонкие волокна, берущие начало от ядер серого бугра. Наружная часть срединного возвышения образована опорными нейроглиальными (эпендимными) волокнами, между к-рыми залегают многочисленные нервные волокна. В этих нервных волокнах и около них наблюдается отложение нейросекреторных гранул. В наружном слое срединного возвышения расположена сеть капилляров, обеспечивающая кровоснабжение аденогипофиза. Эти капилляры образуют петли, поднимающиеся в толщу срединного возвышения навстречу нервным волокнам, которые спускаются к этим капиллярам.
Г. включает ядра, образованные нервными клетками, не обладающими секреторной функцией, и ядра, состоящие из нейросекреторных клеток. Секреторные нервные клетки сконцентрированы гл. обр. непосредственно около стенок III желудочка. По своим структурным особенностям эти клетки напоминают клетки ретикулярной формации (см.). Физиол, данные свидетельствуют о том, что клетки такого типа продуцируют физиологически активные вещества, способствующие выделению тройных гормонов из гипофиза и названные гипоталамическими нейрогормонами (см.).
Нейросекреторные клетки сосредоточиваются в передней области Г., где образуют с каждой стороны надзрительное (nucl. supraopticus) и околожелудочковое (nucl. paraventricularis) ядра. Надзрительное ядро расположено в задне-боковой области от начала зрительного тракта. Оно образовано группой клеток, лежащих вдоль угла, между стенкой III желудочка и дорсальной поверхностью зрительного перекреста. Околожелудочковое ядро состоит из нервных клеток крупных и средних размеров, имеет вид пластинки, залегающей между сводом (fornix) и стенкой III желудочка, начинается в области зрительного перекреста и постепенно поднимается кзади и кверху в наклонном направлении.
Между обоими названными ядрами расположены многочисленные одиночные нейросекреторные клетки или их группы. В околожелудочковом ядре крупные нейросекреторные клетки сосредоточены преимущественно в расширенной задней части (крупноклеточная часть), а в суженной передней части этого ядра преобладают нейроны меньшего размера. Область надзрительного и околожелудочкового ядер характеризуется обильной васкуляризацией. Аксоны нейронов околожелудочкового и надзрительного ядер, образуя гипоталамо-гипофизарный пучок, достигают задней доли гипофиза, где образуют контакты с капиллярами. В задней доле гипофиза нейрогормоны накапливаются и поступают в ток крови. Основной особенностью нейросекреторных клеток является наличие специфических (элементарных) гранул, содержащихся в разном количестве как в области перикарионов, так и в отростках - аксонах и дендритах (см. Гипоталамо-гипофизарная система). Нейросекреторные клетки надзрительного и околожелудочкового ядер сходны между собой по форме и структуре, но допускается определенная дифференцировка; клетки надзрительного ядра вырабатывают преимущественно антидиуретический гормон (см. Вазопрессин), а околожелудочкового - окситоцин (см.). Т. о., Г. образован комплексом нервно-проводниковых и нейросекреторных клеток. В связи с этим регулирующие влияния Г. передаются к эффекторам, в т. ч. и к эндокринным железам, не только с помощью гипоталамических нейрогормонов, переносимых с током крови и, следовательно, действующих гуморально, но и по эфферентным нервным волокнам.
Г. тесно связан с соседними структурами головного мозга проводящими путями. С передним мозгом Г. связан медиальным пучком, волокна к-рого возникают в обонятельной луковице, головке хвостатого ядра, миндалевидном теле и передней части парагиппокампальной извилины (gyrus parahippocampalis).
Г. обладает хорошо развитой и очень сложной системой афферентных и эфферентных путей. Афферентные пути Г. разделяются на шесть групп: 1) медиальный пучок переднего мозга, связывающий перегородку и преоптическую область почти со всеми ядрами Г.; 2) свод, являющийся системой афферентных волокон, соединяющих кору гиппокампа (см.) с Г.; основная часть волокон свода идет к ядрам сосцевидного тела, другая - в перегородку и в латеральную преоптическую область, третья - к другим ядрам Г.; 3) таламо-гипофизарные волокна, соединяющие в основном медиальные и внутрипластиночные ядра таламуса (см.) с Г.; 4) сосцевидно-покрышечный пучок, в к-ром находятся волокна, восходящие из среднего мозга (см.) к Г.; часть этих волокон заканчивается в преоптической области и перегородке; 5) задний продольный пучок (fasciculus longitudinalis dorsalis), несущий импульсы от ствола мозга к Г.; система волокон заднего продольного пучка и сосцевидных тел обеспечивает связь ретикулярной формации среднего мозга с Г. и лимбической системой (см.); 6) паллидо-гипоталамический путь, связывающий стрио-паллидарную систему с Г. Установлены также непрямые мозжечково-гипоталамические связи, оптико-гипоталамические пути, вагосупраоптические связи.
Эфферентные пути Г. подразделяются на три группы: 1) пучки волокон перивентрикулярной системы (fibrae periventriculares), берущие начало в задних гипоталамических ядрах, вначале идут вместе через перивентрикулярную зону; некоторые из них оканчиваются в задне-медиальных таламических ядрах; большинство волокон перивентрикулярной системы идет к нижней части ствола мозга, а также к ретикулярной формации среднего мозга и спинному мозгу (ретикулярный тракт Г.); 2) сосцевидные пучки, берущие начало в ядрах сосцевидного тела Г., разделяются на два пучка: сосцевидно-таламический (fasc. mamillothalamicus), идущий к передним ядрам таламуса, и сосцевидно-покрышечный пучок (fasc. mamillotegmentalis), идущий к ядрам среднего мозга; 3) гипоталамо-гипофизарный тракт - самый короткий, но ясно очерченный пучок аксонов нейронов Г.; эти волокна берут начало в надзрительном и околожелудочковом ядрах и идут через гипофизарную ножку к нейрогипофизу. Большинство функций Г., в особенности контроль висцеральных функций, осуществляется через эти афферентные пути. Кроме афферентных и эфферентных связей, в Г. имеется комиссуральный путь. Благодаря ему медиальные Гипоталамические ядра одной стороны вступают в контакт с медиальными и с латеральными ядрами другой стороны.
Основным источником артериального кровоснабжения ядер Г. являются веточки артериального круга мозга, которые обеспечивают изолированное обильное кровоснабжение отдельных групп ядер Г. Сосуды Г. отличаются высокой проницаемостью для крупномолекулярных белковых соединений. Взаимосвязь между Г. и аденогипофизом осуществляется через сосуды портальной системы, к-рая имеет свои особенности (см. Гипоталамо-гипофизарная система).
Физиология
Г. занимает ведущее положение в осуществлении регуляции многих функций целого организма, и прежде всего постоянства внутренней среды (см. Гомеостаз). Г. - высший вегетативный центр, осуществляющий сложную интеграцию и приспособление функций различных внутренних систем к целостной деятельности организма. Он имеет существенное значение в поддержании оптимального уровня обмена веществ (белкового, углеводного, жирового, водного и минерального) и энергии, в регуляции температурного баланса организма, деятельности пищеварительной, сердечно-сосудистой, выделительной, дыхательной и эндокринной систем. Под контролем Г. находятся такие железы внутренней секреции, как гипофиз, щитовидная, половые, поджелудочная, надпочечники и др.
Регуляция тройных функций гипофиза осуществляется путем выделения гипоталамических нейрогормонов, поступающих в гипофиз через портальную систему сосудов. Между Г. и гипофизом имеется обратная связь (рис. 2), с помощью к-рой регулируется их секреторная функция. Принцип обратной связи (feedback relation) заключается в том, что при увеличении секреции гормонов железами внутренней секреции уменьшается секреция гормонов Г. (см. Нейрогуморальная регуляция). Выделение тройных гормонов гипофиз а приводит к изменению функций эндокринных желез, секрет которых попадает в кровь и в свою очередь может действовать на Г. В Г. обнаружено семь гипоталамических нейрогормонов активирующих и три - ингибирующих выделение тройных гормонов гипофиза. Они широко используются в клинике с целью диагностики заболеваний желез внутренней секреции. Принято считать, что передняя область Г. принимает непосредственное участие в регуляции выделения гонадотропинов. Большинство исследователей считает центром, регулирующим тиреотропную функцию гипофиза, область, расположенную в переднебазальной части Г., ниже околожелудочкового ядра, простирающуюся от надзрительных ядер спереди до аркуатных ядер кзади. Локализация областей, избирательно контролирующих адренокортикотропную функцию гипофиза, изучена недостаточно. Ряд исследователей связывает регуляцию АКТГ с задней областью Г. Венгерская школа Сентаготаи (J. Szentagothai) связывает регуляцию АКТГ с премамиллярной областью. Максимальная концентрация АКТГ - рилизинг-фактора обнаруживается в области медиальной эминенции. Локализация областей Г., участвующих в регуляции остальных тропных гормонов гипофиза, остается неясной. Функциональное обособление и разграничение гипоталамических зон по их участию в контролировании тропных функций гипофиза невозможно провести достаточно четко.
Многочисленные исследования показали, что передняя область Г. оказывает стимулирующее влияние на половое развитие, а задняя область Г.- тормозящее. У больных с патологией гипоталамической области наблюдается нарушение функций половой системы: половая слабость, нарушение менструального цикла. Известно много случаев быстрого полового созревания в результате чрезмерного раздражения опухолью области серого бугра. При адипозогенитальном синдроме, связанном с поражением туберальной области Г., наблюдаются нарушения половой функции.
Г. имеет важное значение в поддержании оптимальной; температуры схемы тела (см. Терморегуляция).
Механизм потери тепла связан с функцией передней области Г. Разрушение задних отделов Г. вызывает снижение температуры тела.
Г. регулирует функцию симпатической и парасимпатической частей вегетативной нервной системы, их координацию. Задняя область Г. участвует в регуляции активности симпатической части в. н. с., а средняя и передняя - парасимпатического отдела, т. к. стимуляция передней и средней областей Г. вызывает парасимпатические реакции (урежение сердцебиений, усиление перистальтики кишечника, тонуса мочевого пузыря и др.), а раздражение задней области вызывает симпатические реакции (учащение сердцебиения и т.д.). Между этими центрами существуют Реципрокные связи. Однако четко разграничить центры в Г. трудно.
Изучение гипоталамического уровня регуляции пищевого поведения показало, что она осуществляется в результате реципрокных взаимодействий двух пищевых центров: латерального и вентро-медиального гипоталамических ядер. Активация нейронов латерального Г. вызывает формирование пищевой мотивации. При двустороннем разрушении этого отдела Г. пищевая мотивация полностью устраняется, и животное может погибнуть от истощения. Повышение активности вентро-медиального ядра Г. снижает уровень пищевой мотивации. При разрушении этого ядра значительно повышается уровень пищевой мотивации, наблюдается гиперфагия, полидипсия и ожирение.
Вазомоторные реакции гипоталамического происхождения тесно связаны с состоянием в. н. с. Различные виды артериальной гипертензии (см. Гипертензия артериальная), развивающиеся после стимуляции Г., обусловлены комбинированным влиянием симпатического отдела в. н. с. и выделением адреналина из надпочечников. Однако в данном случае нельзя исключить также влияние нейрогипофиза, особенно в генезе устойчивой гипертензии, что подтверждается экспериментальными данными, когда артериальная гипертензия, вызванная стимуляцией задней области Г., снижается после электрического разрушения медиальной эминенции. Региональные вазомоторные реакции, развивающиеся после разрушения преоптической области, отличаются от общих вазомоторных реакций, наблюдаемых после стимуляции задней области Г.
Г.- одна из главных структур, участвующих в регуляции смены сна и бодрствования (см. Сон). Клин, исследованиями установлено, что симптом летаргического сна при эпидемическом энцефалите обусловлен повреждением Г. Повреждение Г. вызывало сон и в эксперименте. Для поддержания состояния бодрствования решающее значение имеет задняя область Г. Обширное разрушение средней области Г. приводило к состоянию длительного сна у животных. Нарушение сна в виде нарколепсии объясняется поражением ростральной части ретикулярной формации среднего мозга и Г. Получены экспериментальные данные (П. К. Анохин, 1958), свидетельствующие о том, что сон как результат торможения корковой деятельности развивается в результате высвобождения гипоталамических образований, которые остаются активными в течение всего периода сна.
Г. находится под регулирующим влиянием коры головного мозга. Нейроны коры, получая информацию об исходном состоянии организма и окружающей среды, оказывают нисходящие влияния на все подкорковые структуры, в т. ч. и на центры Г., регулируя уровень их возбуждения. Кора больших полушарий оказывает тормозящее влияние на функции Г. Приобретенные корковые механизмы подавляют многие эмоции и первичные побуждения, формирующиеся с участием Г. Поэтому декортикация нередко приводит к развитию реакции «мнимой ярости» (расширение зрачков, пилоэрекция, тахикардия, повышение внутричерепного давления, саливация и т. д.).
Г. с физиол, точки зрения имеет ряд особенностей, и прежде всего это касается его участия в формировании поведенческих реакций организма, важных для сохранения постоянства внутренней среды. Раздражение Г. приводит к формированию целенаправленного поведения - пищевого, питьевого, полового, агрессивного и т. п. Г. принадлежит главная роль в формировании основных влечений организма (см. Мотивации).

Метаболизм нейронов Г.. избирательно чувствителен к содержанию тех или иных веществ в крови, и при любом изменении их содержания эти клетки приходят в состояние возбуждения. Гипоталамические нейроны чувствительны к малейшим отклонениям pH крови, напряжения углекислоты и кислорода, содержания ионов, особенно калия и натрия и др. Так, в супраоптическом ядре Г. обнаружены клетки, избирательно чувствительные к изменению осмотического давления крови, в вентро-медиальном ядре - содержания глюкозы, в переднем гипоталамусе - половых гормонов. Т. о., клетки Г. выполняют функцию рецепторов, воспринимающих изменение гомеостаза, и обладают способностью трансформировать гуморальные изменения внутренней среды в нервный процесс, биологически окрашенное возбуждение. Центры Г. характеризуются выраженной избирательностью возбуждения в зависимости от различных изменений состава крови (рис. 3). Клетки Г. могут избирательно активироваться не только при изменении определенных констант крови, но и нервными импульсами из соответствующих связанных с данной потребностью органов. Нейроны Г., обладающие избирательной рецепцией по отношению к изменяющимся константам крови, работают по триггерному типу (см. Триггерные механизмы). Возбуждение в этих клетках Г. возникает не сразу, как только изменится какая-либо константа крови, а через определенный промежуток времени, когда возбудимость их повысится до критического уровня. Т. о., клетки мотивационных центров Г. характеризует периодичность работы. Если же изменение константы крови поддерживается длительно, то в этом случае возбудимость нейронов Г. быстро поднимается до критической величины и состояние возбуждения этих нейронов поддерживается на высоком уровне все время, пока существует изменение константы, вызвавшее развитие процесса возбуждения. Постоянная импульсация нейронов Г. устраняется только тогда, когда исчезает вызывавшее ее раздражение, т. е. нормализуется содержание того или другого фактора крови. Функционирование триггерных, механизмов Г. значительно растянуто во времени. Возбуждение одних клеток Г. может возникать периодически через несколько часов, как, напр., при недостатке глюкозы, других - через несколько суток или даже месяцев, как, напр., при изменении содержания половых гормонов. Нейроны Г. не только воспринимают изменения показателей крови, но и трансформируют их в специальный нервный процесс, формирующий поведение организма в окружающей среде, направленное на удовлетворение внутренней потребности.
Обширные связи Г. с другими структурами головного мозга способствуют генерализации возбуждений, возникающих в клетках Г. В первую очередь возбуждение из Г. распространяется на лимбические структуры мозга и через ядра таламуса на передние отделы коры больших полушарий. Зона распространения восходящих активирующих влияний Г. зависит от силы исходного раздражения центров Г. При усилении возбуждения центров Г. активируются аппараты ретикулярной формации. Все эти восходящие активирующие влияния гипоталамических центров, возбужденных внутренней потребностью организма, и определяют возникновение состояния мотивационного возбуждения.
Нисходящие влияния Г. обеспечивают регулирование функций гл. обр. через в. н. с. Но вместе с тем важным компонентом в осуществлении нисходящих влияний Г. являются и гормоны гипофиза. Т. о., как восходящие, так и нисходящие влияния Г. осуществляются нервным и гуморальным путем (см. Нейрогуморальная регуляция). Нисходящим влияниям Г. уделяется большое внимание в связи с концепцией Г. Селье о реакции «стресс» (см. Адаптационный синдром , Стресс). Установлено существование тормозящих влияний различных ядер Г. на моно- и полисинаптические спинальные рефлексы. При раздражении комплекса мамиллярных ядер в некоторых случаях отмечается повышение активности мотонейронов спинного мозга.
Г. находится в непрерывных циклических взаимодействиях с другими отделами подкорки и корой головного мозга. Именно этот механизм лежит в основе участия Г. в эмоциональной деятельности (см. Эмоции). Особое значение центров Г. в деятельности целого организма позволило П. К. Анрхину и К. В. Судакову (1968,1971) высказать предположение о "пейцмекерной" (пейцмекер - пусковой механизм) роли этой структуры мозга в формировании биол, мотиваций. Благодаря тому, что к гипоталамических отделам адресуется нервная и гуморальная сигнализация о различных внутренних потребностях, они и приобретают значение «пейцмекеров» мотивационных возбуждений. Согласно этому представлению, Гипоталамические «пейцмекеры» благодаря восходящим активирующим влияниям определяют энергетическую основу мотивационных возбуждений.
Нейроны мотивационных центров Г. обладают различной хим. спецификой, к-рая определяется избирательным использованием в их метаболизме специальных хим. веществ. И эта хим. специфика Г. сохраняется в восходящих активирующих его влияниях на всех уровнях, обеспечивая качественное биол, своеобразие поведенческих актов. Так, введением адренолитических веществ (аминазин) удается избирательно блокировать механизмы активации коры головного мозга при ноцицептивном раздражении. Активация коры головного мозга при пищевом возбуждении голодных животных избирательно блокируется холинолитическими препаратами. Нейротропные вещества со специфическим механизмом действия благодаря существованию гетерохим. организации гипоталамических центров могут избирательно блокировать различные механизмы Г., участвующие в формировании таких состояний организма, как голод, страх, жажда и т. д.
Методы исследования
Электроэнцефалографический метод. По результатам электроэнцефалографического исследования поражения (см. Электроэнцефалография) могут быть подразделены на четыре группы: первая группа - отсутствие отклонений или минимальные отклонения от нормальной ЭЭГ; вторая группа - резкое снижение альфа-ритма вплоть до его исчезновения; третья группа - появление на ЭЭГ тэта-ритма, особенно в связи с повторными афферентными раздражениями; четвертая группа - приступообразные нарушения ЭЭГ в виде появления изменений, характерных для сна; этот тип ЭЭГ характеризует диэнцефальную эпилепсию. При описанных выше синдромах сравнительная оценка ЭЭГ не обнаруживает специфичности.
Плетизмографические исследования (см. Плетизмография) выявляют широкий спектр изменений - от состояния вегетативной сосудистой неустойчивости и парадоксальной реакции до полной арефлексии (см.), что соответствует степени выраженности функциональных или органических поражений ядер Г. При исследованиях в. н.д. с помощью двигательного метода с речевым подкреплением было установлено, что при всех формах патологии Г. взаимодействие между корой и подкоркой резко снижено.
У больных с поражением Г. независимо от его причины (опухоль, воспаление и др.) может увеличиваться содержание катехоламинов и гистамина в крови, увеличивается альфа-глобулинов фракция и снижается бета-глобулиновая фракция, изменяется уровень экскреции 17-кетостероидов. При различных формах поражения Г. отчетливо проявляются нарушения кожной температуры и потоотделения.
Патология
В гипоталамусе возникают как функциональные нарушения, так и необратимые изменения его ядер. В первую очередь следует отметить возможность различной степени поражения ядер (преимущественно надзрительного и околожелудочкового) при заболеваниях желез внутренней секреции.
Травмы мозга, приводящие к перераспределению церебральной жидкости, также могут вызывать изменения в гипоталамических ядрах, расположенных вблизи эпендимы дна III желудочка.
Патоморфологически эти изменения касаются прежде всего нейронов и особенно четко выявляются при окраске по Нисслю (см. Ниссля метод) и методом Гомори. Они выражаются явлениями тигролиза, нейронофагии, вакуолизацией протоплазмы, образованием клеток теней. Вследствие повышенной проницаемости стенок сосудов при инфекциях и интоксикациях Гипоталамические ядра могут подвергаться патогенным воздействиям токсинов и хим. продуктов, циркулирующих в крови. Особенно опасны нейровирусные инфекции. Наиболее часто встречающиеся воспалительные процессы Г.- базальный менингит туберкулезного происхождения и сифилис. К редким формам поражения Г. относятся гранулематозные воспаления (болезнь Бека), лимфогранулематоз, лейкемия, а также сосудистые аневризмы различного происхождения. Из опухолей Г. наиболее часто встречаются различного вида глиомы, определяемые как астроцитомы; краниофарингеомы, эктопические пинеаломы и тератомы, а также супраселлярные аденомы гипофиза, расположенные над турецким седлом, менингиомы и кисты.
Клинические проявления нарушений функции гипоталамуса
При поражении Г. выделяют следующие основные синдромы.
1. Нейро-эндокринный , проявляющийся ожирением с характерным перераспределением подкожной жировой клетчатки (лунообразное лицо, толстая шея и туловище, худые конечности), остеопорозом с наклонностью к кифозу позвоночника, болями в спине и пояснице, нарушением половой функции (ранняя аменорея у женщин и импотенция у мужчин), ростом волос на лице и туловище у женщин и подростков, гиперпигментацией кожи, особенно в местах складок, наличием багровых атрофических полос на животе и бедрах (striae distensae), артериальной гипертензией, периодическими отеками, общей слабостью и повышенной утомляемостью. Разновидностью указанного синдрома является Иценко - Кушинга болезнь (см.).
Другие проявления нейро-эндокринного синдрома - диабет несахарный (см.), Гипофизарная кахексия (см.), адипозо-генитальная дистрофия (см.) и др.
2. Нейродистрофический синдром характеризуется изменением солевого обмена, деструктивными изменениями в коже и мышцах, сопровождающимися отеками и атрофией кожн, нейромиозитами, периодически наступающими внутрисуставными отеками; кожа сухая, шелушащаяся с полосами растяжений, наблюдаются зуд, сыпи. Отмечается также остеомаляция, кальцификация, склерозирование костей, образование язв, пролежней, кровотечения по ходу жел.-киш. тракта и в паренхиме легких, преходящие отеки сетчатки глаза.
3. Вегетативно-сосудистый синдром характеризуется расширением мелких вен на лице и теле, повышенной ломкостью сосудов, наклонностью к геморрагиям, высокой проницаемостью стенок сосудов, различными вегетативно-сосудистыми пароксизмами, в т. ч. мигренями, сопровождающимися повышением или понижением АД.
4. Невротический синдром проявляется своеобразными истерическими реакциями и психопатол, состояниями, а также нарушениями бодрствования и сна.
Перечисленные синдромы могут проявляться как при функциональных нарушениях, так и при органических поражениях ядер Г. Если вегетативно-сосудистый синдром отмечается при функциональных изменениях, то нейродистрофический - при тяжелых органических поражениях ядер средней области Г., иногда передней и задней его областей. Нейро-эндокринный синдром проявляется вначале как результат функциональных нарушений ядер передней области Г., в дальнейшем присоединяются органические поражения упомянутых ядер.
Лечение
При патологии гипоталамической области применяются три вида лечения.
1. Рентгенотерапия малыми дозами в пределах (50 р) 6-8 сеансов на область Г. при воспалительной природе поражения или наличии резко выраженного аллергического состояния. При хорошей выделительной функции почек облучение следует сопровождать назначением малых доз диуретиков. Рентгенотерапия показана при выраженном вегетативно-сосудистом синдроме, при нейро-эндокринном, в начальной стадии его развития.
2. Гормонотерапия в виде моно-терапии или в сочетании с рентгенотерапией. Применение кортизона, преднизолона или их производных, а также АКТГ должно сопровождаться тщательным наблюдением за гормональной функцией надпочечников. Применяются также препараты половых гормонов щитовидной железы, проводятся попытки применения ри лизинг-гормонов.
3. Введение методом ионогальванизации в слизистую оболочку носа различных хим. веществ при минимальной силе тока 0,3-0,5 а; длительность процедуры 10-20 мин. Обычно проводится до 30 сеансов. При ионогальванйзащш используют 2% р-р хлорида кальция, 2% р-р витамина B1, 0,25% р-р димедрола, р-р эрготамина или фенамина. Ионогальванизация несовместима с рентгенотерапией. В ряде случаев применяют средства, снижающие внутричерепное давление, действующие на процессы торможения или возбуждения в коре и подкорке (фенобарбитал, бромиды, кофеин, фенамин, эфедрин). Во всех случаях необходим тщательный индивидуальный выбор форм лечения.
Оперативное лечение проводится при опухолях Г. по общепринятым методам операций на головном мозге (см.).
Библиография: Баклаваджян О. Г. Гипоталамус, в кн.: Общая и частная физиол. нервн. системы, под ред. П. К. Кос-тюка и др., с. 362, Л., 1969; Г р а щ e н-к о в Н. И. Подбугорье (гипоталамическая область), в кн.: Физиол, и патол, диэнцефальной области головного мозга, под ред. Н. И. Гращенкова и Г. Н. Кассиля, с. 5, М., 1963, библиогр.; о н ж е, Гипоталамус, его роль в физиологии и патологии, М., 1964, библиогр.; С e н т а г о-тай Я. и др. Гипоталамическая регуляция передней части гипофиза, пер. с англ., Будапешт, 1965; Ш а де Дж. и Ф о р д О. Основы неврологии, пер. с англ., М., 1976, библиогр.; H e s s W. R. Hypothalamus und Thalamus, experimen-tal-dokumente, Stuttgart, 1956, Bibliogr.; The hypothalamus, ed. by L. Martini a. o., N. Y.-L., 1970; Schreider Y. The hypothalamo-hypophysial system, Prague, 1963, bibliogr.
B. H. Бабичев, С. А. Осиповский.
Роль гипоталамуса
Гипоталамус, или подбугровая область промежуточного мозга, является высшим центром интеграции и регуляции вегетативных функций организма. Он принимает участие в корреляции различных соматических функций, регуляции работы желудочно-кишечного тракта, сна и бодрствования, водно-солевого, жирового и углеводного обмена, поддержания температуры тела и гомеостаза. Одна из наиболее важных функций гипоталамуса связана с регуляцией деятельности эндокринной системы организма. Разнообразие функции гипоталамуса обусловлено сложностью его морфологического строения и обилием связей с различными отделами нервной системы, органами чувств, внутренними органами и внутренней средой организма. Строение гипоталамуса . Гипоталамус относится к филогенетически древним образованиям мозга и хорошо развит уже у низших позвоночных. Он образует дно третьего желудочка и лежит между перекрестом зрительных нервов и задним краем маммилярных тел. В состав гипоталамуса входит серый бугор, срединное возвышение, воронка и задняя или нервная доля гипофиза. Спереди он граничит с преоптической областью, которую отдельные авторы также включают в систему подбугорья. Гипоталамус развивается в ранний период эмбриогенеза из переднего мозгового пузыря. В процессе развития головного мозга, после обособления больших полушарий, передний мозговой пузырь дает начало межуточному мозгу, а его полость превращается в третий желудочек. В дне этого желудочка путем выпячивания образуется мозговая воронка, дистальный конец которой превращается в заднюю долю гипофиза. Основание воронки значительно утолщается и дает начало серому бугру. В ка-удальной части образуются парные маммилярные тела. Боковые стенки третьего желудочка образуют зрительные бугры, связанные с большими полушариями головного мозга. Центральное серое вещество гипоталамуса без резкой границы переходит в центральное серое вещество среднего мозга. Нервные клетки в гипоталамусе собраны в более или менее обособленные группы или ядра, которые занимают в нем определенное место и состоят из различных по своему строению невронов. Разнообразие нейрального состава ядер гипоталамуса обусловлено их функциональной дифференциройкой. В процессе эволюции ряда животных число и структура гипоталамических ядер подверглись значительным изменениям. У кролика, например, насчитывают 30, а у собаки 15 пар ядер. В литературе пока отсутствует единая номенклатура гипоталамических ядер. Пинес и Майман выделяют в гипоталамусе передний, средний и задний отделы. В каждом отделе они различают следующие ядра. Передний отдел : 1) супрахиазматическое; 2) супраоптическое (передние, латеральные и медиальные отделы); 3) пара-вентрикулярное. Средний отдел : 1) супраоптическое (задние отделы); 2) ту-беральные (верхние, средние и нижние); 3) паллидо-инфунди-булярное; 4) маммило-инфундибулярное. Задний отдел : 1) маммило-йнфундибулярные; 2) ядра маммилярных тел (внутреннее, наружное, вставочное); 3) супра-маммилярные. Филогенетически наиболее древними образованиями гипоталамуса являются паравентрикулярное и супраоптическое ядра. Они гомологичны преоптическим ядрам низших позвоночных. У млекопитающих суираоптическое ядро лежит в переднем гипоталамусе над хиазмой (рис. 1) и проходит в дорсолатеральном направлении от зрительного перекреста до середины серого, бугра.
Рис. 72. Сагиттальный срез мозга млекопитающего (по Кларку): 1 -- гипофиз; 2 -- супрахиазматическое ядро; 3 -- супраоптическое ядро; 4 -- паравентрикулярное ядро; 5 -- ядра мамми-лярной области; 6 -- ядра преоптической области; 7 -- свод; 8-- Stria terminilis -- хиазма; 10--передняя комиссура.
У многих животных оно подразделяется на отдельные группы нейронов, соединенных между собой клеточными мостиками. Не менее характерной группировкой клеток гипоталамуса является паравентрикулярное ядро, расположенное под передней коммисурой в стенке третьего желудочка. Паравентрикулярное ядро развивается из того же самого клеточного материала, что и супраоптическое ядро. В строении клеток этих ядер обнаруживается значительное сходство. Они имеют округлую, грушевидную или удлиненную форму и от нейронов других ядер гипоталамуса и центрального серого вещества отличаются значительно более крупными размерами.

Рис. 2. Схема васкулярных связей гипоталамуса и гипофиза (по Алешину): / -- супраоптическое ядро; 2 -- паравентрикулярное ядро; 3 -- туберальные ядра; 4 -- маммилярные ядра; 5 -- первичная капиллярная сеть в срединном возвышении; 6 -- сосудистые клубочки первичной капиллярной сети; 7 -- вены портальной системы; 8 -- передняя доля гипофиза; 9 -- средняя доля гипофиза; 10 -- задняя доля гипофиза; // -- вторичная капиллярная система в передней доле гипофиза; 12 -- капилляры задней доли гипофиза; 13 -- гипоталамо-гипо-физарный тракт.
Васкуляризация гипоталамуса . Гипоталамическая область характеризуется обильным кровоснабжением. Наибольшей васкуляризацией отличается паравентрикулярное и супраоптическое ядра, в которых каждая клетка связана с 2--3 капиллярами. Здесь на площадь 1 мм 2 приходится до 2650 капилляров. Электронно-микроскопические исследования показали, что в местах контакта между телом нейрона и сильно утонченной базальной мембраной эндотелия капилляров часто совсем нет глиальной прослойки. Вследствие этого сосуды обладают очень хорошей проницаемостью даже для высокомолекулярных белковых соединений. Из крови в клетки супраоптических и паравентрикулярных ядер легко поступают питательные вещества, гормоны и другие химические соединения. Гипоталамические образования поэтому обладают высокой чувствительностью к отклонениям в составе гуморальной среды организма и отвечают на них изменением физиологической активности.
Важнейшее значение в механизме гипоталамической регуляции гормональной функции гипофиза имеет общность их васкуляризации. Между гипоталамусом и передней долей гипофиза существует специальная система кровообращения, получившая название воротной, или портальной, системы сосудов гипофиза. Она состоит из артериол, которые берут начало от артерий виллизиевого круга. Артериолы проникают в срединное возвышение серого бугра и здесь распадаются на большое количество капилляров. В срединном возвышении клубочки и петли этих капилляров вступают в тесный контакт с окончаниями нервных волокон клеток нейросекреторных ядер гипоталамуса и образуют с ними так называемые вазоневральные синапсы (рис. 2). Первичные капилляры в сером бугре собираются в портальные вены, которые по гипофизарной ножке идут в переднюю долю гипофиза, и в его паренхиме распадаются в густую сеть синуооидных капилляров (вторичная капиллярная сеть). В заднюю долю гипофиза сосуды портальной системы не проникают, и кровь в нее поступает из других источников. Движение крови по портальной системе от гипоталамуса к гипофизу происходит в результате сокращения стенок сосудов. В переднюю долю гипофиза кровь поступает еще по средней и задней гипофизарным артериям, а также по сосудистым анастомозам из нейрогипофиза.
Связи гипоталамуса . Гипоталамическая область имеет обширные связи с различными отделами центральной нервной системы, в том числе с ретикулярной формацией ствола мозга, гипофизом и т. д. Среди проводящих путей гипоталамуса различают эфферентные, афферентные и внутригипоталамические связи.
Эфферентные пути от гипоталамуса идут к таламусу (гипо-таламо-таламический путь), покрышке (маммило-тегментальный путь), от всех ядер гипоталамуса к нижележащим симпатическим образованиям и узлам (диффузные нисходящие связи), от гипоталамуса к гипофизу (гипоталамо-гипофизарный тракт). В гипоталамо-гипофизарных путях выделяют супраоптикогипо-физарный, а также туберо-гипофизарный пути. Первый путь образован большим количеством (до 100000) аксонов клеток су-праоптического и паравентрикулярного ядер, которые по ножке гипофиза поступают в заднюю долю гипофиза. Эти волокна проходят во внешнем слое срединного возвышения и не проникают из задней в переднюю долю гипофиза (см. рис. 2).
В регуляции функций эндокринной системы особое значение имеет тубероинфундибулярный путь, который передает эфферен-тацию из гипоталамуса в переднюю долю гипофиза. Волокна этого пути прослеживаются до срединного возвышения, где их окончания с петлями и клубочками первичных капилляров портальной системы образуют рассмотренные выше вазоневральные синапсы.
Афферентные пути к ядрам гипоталамуса идут от таламуса, лобных долей, гиппокампа, зрительного бугра,- амигдалоидного комплекса, миндалин, экстрапирамидной системы и ретикулярной формации ствола мозга.
Ретикулярной формации приписывается большое значение в регуляции функции гипоталамуса и эндокринной системы. Исследованиями Грина, Русселя и др. установлено, что ядра гипоталамуса находятся в тесной анатомической и.функциональной связи с ретикулярной формацией. Последняя образована сложным комплексом невронов различной величины, которые диф-фузно рассеяны в стволе мозга. Для отростков клеток ретикулярной формации характерно наличие большого количества коллатералей, через посредство которых один аксон может вступать в функциональные связи с множеством нервных клеток (до 20 000). Ретикулярная формация, как впервые было установлено Мэгуном и Мурицци, оказывает общее активирующее действие на различные отделы мозга и состоит из восходящей и- нисходящей систем. Волокна восходящей системы от каудальных частей продолговатого мозга, варолиева моста и среднего мозга проецируются на различные участки коры больших полушарий; нисходящие же волокна связывают ретикулярную формацию с системой спинного мозга." В ретикулярную формацию проецируется огромное количество волокон от ядерных образований ствола мозга, рецепторов внутренних органов, от аппаратов зрения, слуха и проводников чувствительности. Ретикулярная формация отличается высокой чувствительностью к изменениям гуморальной среды организма. На воздействие гормонов и различных химических соединений она быстро отвечает изменением своей физиологической активности.
Гипоталамус получает восходящие волокна преимущественно от ретикулярной формации среднего мозга. Через покрышку среднего мозга и задний отдел гипоталамуса эти волокна достигают серого бугра. На этой морфологической основе осуще-ствляется функциональная связь между ретикулярной формацией, гипоталамусом и эндокринными железами. Ретикулярная формация среднего мозга передает импульсы через гипоталамус к эндокринным железам организма и оказывает активизирующее действие на ядра подбугорья.
Нейросекреторная деятельность гипоталамуса . Нейроны отдельных ядер гипоталамуса проявляют способность к секреторной деятельности (нейрокринии) и вырабатывают особые вещества (нейросекреты), которые играют важную роль в регуляции функции эндокринной системы. Пионерами в изучении гипотала-мической нейросекреции являются Шаррер и Гаупп, которые еще в 1933 г. обнаружили в клетках переднего гипоталамуса гранулы и капельки нейросекрета. Последующими исследованиями было установлено широкое распространение явлений нейросекреции не только у позвоночных, но и беспозвоночных животных.
У рыб нейросекреторная функция характерна для преоптического ядра передней области гипоталамуса. У высших позвоночных и человека эти свойства в наиболее отчетливой форме выражены в клетках паравентрикулярного и супраоптического ядер. Нейросекреция характерна и для нейронов ядер серого бугра, но в них она не получает такого яркого морфологического выражения, как в паравентрикулярном и супраоптическом ядрах. гипоталамус соматический гомеостаз нейросекреторный
Нейросекреторные невроны гипоталамуса одновременно сочетают в себе свойства нервных и железистых клеток. Они относятся к мультиполярным невронам с относительно крупным ядром и содержат нейрофибриллы, хорошо развитую систему эргастоплазмы (субстанцию Ниссля) с рибосомами и общие для всех клеток органоиды.
Нейросекреторные процессы у высших животных наиболее полно изучены в клетках супраоптического и паравентрикулярного ядер. Нейросекрет представлен гранулярными образованиями, обладающими известным постоянством своего строения у всех животных. Гранулы имеют вид гомогенных шариков и пузырьков, окруженных мембраной. В зависимости от специфического отношения к красителям различают гомориположительный и гомориотрицательный нейросекрет. Первый хорошо окрашивается хромовоквасцовым гематоксилином по Гомори в темно-синий цвет; гомориотрицательное или оксифильное нейросекре-торное вещество при окраске по Гомори красится флоксином в розовый цвет.
Химическая природа нейросекрета окончательно не выяснена. Гомориположительное вещество обладает относительно высокой химической устойчивостью и является сложным белково-полиса-харидо-липидным соединением. Гомориотрицательный нейросекрет является относительно простым протеином, богатым аминокислотами с сульфгидрильными и дисульфидными группами. Этот вид нейросекрета наиболее широко распространен у беспозвоночных животных.
Первичный биосинтез нейросекрета в клетках происходит в приядерной зоне цитоплазмы. В перинуклеарной области он появляется в виде мелких пылевидных зернышек, которые затем распространяются по всей клетке (рис. 3). Образование нейросекрета связано с уменьшением размеров ядра и ядрышка, а также заметной редукцией субстанции Ниссля. Электронномикро-скопические исследования показывают, что в синтезе нейросекрета главную роль играет эргастоплазма с ее системой рибосом и аппарат Гольджи (Шаррер и др.). Считают, что образование нейросекрета в клетках ядер гипоталамуса протекает по апокриновому, меракриновому и голокриновому типам (Поленов).
Интенсивность синтеза секрета и выведения его из клеток меняется в зависимости от времени года, условий температурного и светового режимов, физиологического состояния организма, стадии полового цикла и т. д. При дегидратации организма,

Рис. 74. Последовательные стадии образования нейро-секрета в клетках гипоталамуса (по Шарреру): 1 -- тельца Ниссля; 2 -- гранулы секрета; 3 -- аксон; 4 -- ядро; 5 -- базофильная цитоплазма.
например, в клетках паравентрикулярного и супраоптического ядер значительно уменьшается содержание нейросекреторного вещества.
Возрастные изменения гипоталамической нейросекреции изучены еще недостаточно. Образование нейросекрета в супраопти-ческих и паравентрикулярных ядрах гипоталамуса начинается уже в ранний период индивидуального развития животных. У крупного рогатого скота, например, в ядрах подбугорья нейро-секрет обнаруживается у 3-месячных зародышей. У кур первые признаки нейросекреции в супраоптических ядрах обнаруживаются в ранний период инкубации. По данным Денисьевского, образование нейросекрета в клетках паравентрикулярных ядер зародыша уток начинается на 17-й день инкубации. У морских свинок нейросекреторный процесс начинается на 21--28-й день после рождения. У многих животных образование нейросекрета наступает в невронах супраоптического ядра раньше, чем в пара-вентрикулярном. В ранний период развития в гипоталамусе содержится меньше нейросекрета, чем у взрослых животных.
Нейросекрет оказывает физиологическое действие через гуморальную среду организма. В связи с этим значительный интерес представляет вопрос о путях его выведения из нейронов ядер гипоталамуса. Микроскопическими исследованиями установлено, что из невронов супраоптических и паравентрикулярных ядер гранулы гомориположительного нейросекрета по длинным аксонам гипоталамо-гипофизарного тракта токами аксоплазмы смещаются в заднюю долю гипофиза. По мере передвижения от места образования до окончания нервных отростков изменяются химические и тинкториальные свойства нейросекрета. По данным отдельных исследователей, нейросекрет может синтезироваться и в терминалях аксонов, лежащих в задней доле гипофиза (Ди-пен). Задняя доля гипофиза является, таким образом, резервуаром для гипоталамического гомориположительного нейросекрета. Отсюда нейросекрет поступает в кровеносное русло. Часть нейро-секреторного вещества может попадать в кровеносное русло и через ликвор третьего желудочка. Кроме того, нейросекреторные клетки посылают свои аксоны к обонятельным ядрам и эпендиме боковых желудочков переднего мозга. Депонированный в задней доле гипофиза, гомориположительный нейросекрет является носителем физиологически высокоактивных гормонов вазопрессина и окситоцина. По мнению некоторых авторов,_окситоцин образуется в паравентрикулярном, а вазопресин в супраоптическом ядрах. Другие же исследователи считают, что, в зависимости от физиологического состояния организма, клетки одного и того же ядра могут синтезировать оба гормона.

Рис. 75. Портальная система сосудов гипофиза у птиц (по Глису): 1 -- передняя доля гипофиза; 2 -- задняя доля гипофиза; 3 -- срединное возвышение.
Гипоталамический нейросекрет поступает также в переднюю долю гипофиза и контролирует ее гормональную функцию. Однако сюда он поступает из подбугорья не по нервным окончаниям, а с током крови, циркулирующей по сосудам портальной системы. Гуморальный путь тра-нспортировки вырабатываемых клетками гипоталамуса веществ в переднюю долю гипофиза доказывается прямыми экспериментами. У уток, например, портальные вены из гипоталамуса в аденогипофиз проходят обособленно от гипофизарной ножки (рис. 4). Бенуа и Ассенмахер указывают, что перерезка только одной ножки не оказывает существенного влияния на гормональную функцию гипофиза. Перерезка же портальной системы сосудов, при сохранении целостности гипофизной ножки, приводит к угнетению физиологической активности передней доли гипофиза. Показано также, что кровь, взятая из портальных сосудов, стимулирует гипофиз, тогда как кровь из сонной артерии этими свойствами не обладает. Эти эксперименты показывают, что из гипоталамуса в переднюю долю гипофиза действительно поступают вещества, которые активируют его гормональную деятельность. Однако природа этих веществ изучена еще недостаточно.

Рис. 5. Пер. в. с. -- передние воротные сосуды; Зад. в. с. --задние воротные сосуды; МТ -- маммилярпос тело; X -- хиазма; СО -- супраоптическое ядро; ПДГ -- передняя доля гипофиза; ЗДГ -- задняя доля гипофиза
Микроскопическими исследованиями установлено, что во внутреннем слое срединного возвышения амиэлиновые волокна ту-бероинфундибулярного пучка, берущие начало от клеток ядер серого бугра, своими окончаниями вступают в связь с расположенными здесь короткими петлями и клубочками первичных капилляров портальной системы (см. рис. 2).
В этих вазоневральных синапсах отростки нервных клеток гипоталамуса отдают в кровь портальной системы секреторное вещество, которое поступает затем в паренхиму передней доли гипофиза. Этот нейросекрет, выделяемый ядрами серого бугра, однако, не окрашивается по Гомори. Отдельные исследователи отождествляли его с медиаторами нервных клеток (ацетилхоли-ном и норадреналином). В настоящее время эта теория уже почти совсем не имеет сторонников. Исследования показали, что нейро-секрсторное вещество от обычных метаболитов нервных клеток отличается более высокой энзиматической устойчивостью и способностью оказывать физиологическое действие на значительном расстоянии от места своего образования в гипоталамусе. Значительный интерес представляет тот факт, что при электролитическом повреждении отдельных участков срединного возвышения в передней доле гипофиза подавляется образование и выведение в кровь не всех, а только какого-либо одного гормона, активирующего функцию строго определенной периферической эндокринной железы (рис. 5). На этом основании считают, что отдельные нервные волокна от различных клеток гипоталамуса несут в срединное возвышение и отдают в кровь портальной системы не одно, а несколько особых веществ или нейрогуморов (Каррато и др.), которые реализуют различные функции передней доли гипофиза (гонадотропную, тиреотропную и адренокор-тикогропную). Возможно, различные нейрогуморы сосудами портальной системы проецируются на определенные зоны паренхимы передней доли гипофиза.
Как уже отмечалось, значение гомориположительного нейро-секрета в регуляции гормональной функции гипофиза изучено еще недостаточно. Обычными методами окрашивания его не удается выявить в туберо-гипофизарном пути и в крови портальной системы. Вместе с тем вокруг капилляров, в их эндотелии и между секреторными клетками передней доли гипофиза часто накапливается значительное количество гранул, которые проявляют такие же реакции, как и гипоталамический неиросекрет. При избыточном введении в организм поваренной соли происходит быстрое выведение нейросекрета из задней доли гипофиза, супраоптического и паравентрикулярного ядер. В этих условиях нейросекретом обогащаются и секреторные клетки передней доли гипофиза (Войткевич и др.). Отдельные исследователи допускают, что в осуществлении связи гипоталамуса с гипофизом важную роль могут играть вазопресин и окситоцин, носителями которых является неиросекрет паравентрикулярного и супраоптического ядер (Мартини и др.). Следует, однако, отметить, что методом хроматографии Саффрон удалось отделить в экстрактах задней доли гипофиза от вазопресина и окситоцина вещество, которое активирует адренокортикотропную функцию передней доли гипофиза.
Приведенные данные показывают, что вопрос о природе нейросекрета, ответственного за регуляцию функции гипофиза, еще нуждается в дальнейшей разработке. Тем не менее, обширный фактический материал указывает на ведущее значение в гнпота-ламическом контроле функции эндокринной системы васкулярных связей. Срединное возвышение серого бугра гипоталамуса и является тем участком, во внутреннем слое которого через посредство вазоневральных синапсов осуществляется передача влияний с гипоталамуса на переднюю долю гипофиза.
Значение гипоталамуса в регуляции функции эндокринной системы . Гипоталамус принимает участие в нервной и гуморальной регуляции физиологических функций организма. Особенно велико его значение в контроле гормональной деятельности эндокринной системы. Прежде всего гипоталамус сам продуцирует вещества, которые гуморальным путем влияют на отдельные функции организма. Уже отмечалось, что нейроны супраоптического и паравентрикулярного ядер подбугорья синтезируют неиросекрет, который перемещается по нервным отросткам гипоталамо-гипофизарного тракта и аккумулируется в задней доле гипофиза. Этот нейросекрет является носителем физиологически высокоактивных гормонов вазопресина и окситоцина.
Клинические наблюдения и многочисленные экспериментальные исследования последних лет показывают, что гипоталамус оказывает доминирующее влияние на гормональную деятельность передней доли гипофиза и через нее на многие периферические железы внутренней секреции. Этот вывод основан прежде всего на экспериментах по нарушению анатомической связи между гипоталамусом и гипофизом. Так, при перерезке у кроликов (Гаррис), кур (Ширм и Налбандон), уток (Бенуа и Ассен-махер) и других животных гипофизарной ножки резко понижается выделение гипофизом в кровь кринотропных гормонов, активирующих функцию половых желез, коры надпочечников и щитовидной железы. При нарушении связи между гипофизом и гипоталамусом периферические железы переходят в состояние физиологической депрессии. Эта операция особенно сильно отражается на функциональном состоянии половых желез. Если после перерезки ножки портальные сосуды регенерируют и восстанавливается транспортировка из гипоталамуса нейросекрста, то тогда опять нормализуется работа передней доли гипофиза и периферических желез.
Представления о механизмах передачи регулирующих влияний с гипоталамуса на гипофиз за короткую историю разработки этой важной проблемы современной эндокринологии претерпели существенные изменения. На первых этапах ее разработки многие исследователи считали, что влияние гипоталамуса на гипофиз осуществляется через посредство парасимпатической и симпатической нервной системы. Поскольку, однако, прямая нервная связь между ними, по-видимому, отсутствует, то Шаррер уже давно высказал предположение, что гипоталамическая регуляция гормональных функций гипофиза осуществляется преимущественно гуморальным путем при участии нейросекрета. Это положение в дальнейшем подтвердилось не только в опытах с перерезкой портальных сосудов, но и в экспериментах по трансплантации гипофиза в различные органы. При пересадке его гипофизэктомированным животным в почку или в височную долю (Гаррис и Якобсон) гипофиз приживляется, васкуляризируется, но в этих условиях угнетается его гормональная деятельность. Если же этот гипофиз затем подсадить в область срединного возвышения, то после врастания портальных сосудов его гормональная деятельность быстро восстанавливается. Такой же результат получен при совместной инкубации вне организма гипофиза с фрагментами гипоталамуса или при добавлении в культуру экстракта из срединного возвышения подбугорья.
Многочисленные экспериментальные исследования подтверждают, что гипоталамический контроль гормональной функции гипофиза действительно осуществляется через кровь уже рассмотренными прежде нейрогуморами (реализующими факторами). В экспериментальных условиях на функцию передней доли гипофиза может оказывать влияние и суммарный экстракт из нейрогипофиза. На этом основании отдельные исследователи допускают, как это видно на схеме Поленова (таблица I), возможность действия на гормонопоэз передней доли гипофиза и нейросекрета, который поступает в кровеносное русло из нейрогипофиза.
Значительный интерес представляет вопрос о локализации в гипоталамусе участков, ответственных за регуляцию различных гормональных функций гипофиза. При его разработке в настоящее время используются различные приемы. Наиболее широкое применение получил метод точечной электрокаугуляции гипоталамуса, осуществляемой с помощью стереотаксического аппарата, позволяющего производить строго координированные передвижения электродов. Следует, однако, отметить, что и применение стереотаксической техники не устраняет затруднений в решении поставленного вопроса о топографической локализации в гипоталамусе различных зон, регулирующих отдельные функции гипофиза, так как входящие в его состав клеточные компоненты находятся в сложных морфологических и функциональных взаимоотношениях между собой и другими отделами нервной системы. Поэтому повреждение одного участка неизбежно приводит к морфологическим и функциональным нарушениям других компонентов системы. Кроме того, в строении и функциональной дифференцировке отдельных частей гипоталамуса наблюдаются и видовые различия. Вследствие этого полученные различными исследователями данные о значении отдельных участков гипоталамуса в регуляции эндокринных функций организма порою носят противоречивый характер. В настоящее время вполне определенно можно говорить лишь о том, что гипоталамус контролирует гонадотропную, тиреотропную и аденокортико-тропную функции передней доли гипофиза. Для осуществления этих функций, по Бенуа, например, необходима целостность зоны переднего гипоталамуса, расположенной под паравентри-кулярным ядром на границе с преоптическим и туберальным участками.
Рассмотренные выше данные показывают, что гипоталамус и гипофиз в морфологическом и функциональном отношении образуют единую гипоталамо-гипофизарную систему, в которой нервные импульсы переключаются на гуморальные. Значительный интерес, представляет вопрос о механизме работы этого своеобразного пульта управления эндокринными функциями организма. Обширный материал экспериментальных исследований позволяет рассматривать гипоталамус, гипофиз и периферические железы (железы-мишени) как звенья единой системы, функциональная деятельность которой подчиняется принципу обратных связей с самонастройкой на оптимальный для данных условий жизни организма режим работы.
Разработке этих вопросов в свое время много внимания уделял М. М. Завадовский. Давно известно, что избыток в крови гормонов желез-мишеней автоматически приводит к угнетению, а их недостаток -- к стимуляции соответствующих тронных функций передней доли гипофиза. Причем угнетение тронной функции наступает в результате повышения концентрации в крови гормона железы-мишени, при некоторых условиях, по-видимому, может осуществляться и непосредственно через гипофиз. Обратный же механизм регуляции, т. е. стимуляция тропной функции гипофиза понижением содержания в крови гормона железы-мишени, осуществляется при обязательном участии гипоталамуса. Изменение уровня гормона в крови является, таким образом, сигналом, который воспринимается клетками соответствующих ядер гипоталамуса.
При описании васкуляризации гипоталамуса уже отмечалось, что особенности строения стенок капилляров и их проницаемости для сложных химических соединений обеспечивают высокую чувствительность нейронов подбугорья к гормонам. Факт непосредственного действия на нейроны гормонов доказывается многочисленными экспериментами подсадки в соответствующие зоны гипоталамуса ткани эндокринных желез или аппликации синтетическими гормонами. Например, имплантация с помощью стереотаксического аппарата кристаллов полового гормона угнетает гонадотропную функцию гипофиза и физиологическую активность половой железы. Сходный результат дает и трансплантация кусочков яичника. Таким образом, через посредство гормонов гипоталамус получает информацию об уровне активности желез-мишеней и посылает в гипофиз сигналы, в ответ на которые последний через продукцию соответствующих тройных гормонов устраняет неблагоприятные для организма отклонения в функции эндокринной системы. Экспериментальные исследования вместе с тем показывают, что в некоторых случаях афферентная сигнализация от желез-мишеней к гипоталамусу может передаваться и нервнопроводниковым путем. Настройка рассмотренной системы обратных связей носит динамический характер и изменяется прежде всего в различные периоды онтогенеза.
В регуляции функций эндокринных желез принимают также участие внегипоталамические центры нервной системы и прежде всего ретикулярная формация. Хотя разработка этого вопроса находится еще в начальной стадии, тем не менее уже теперь имеются многочисленные доказательства ее участия в контроле гормональной активности отдельных эндокринных желез. Эксперименты показывают, что при блокировании фармакологическими средствами, частичном повреждении или раздражении ретикулярной формации электрическим током наступают значительные изменения в уровне гормональной активности отдельных эндокринных желез.
Ретикулярной формации приписывается большое значение в механизме передачи к эндокринным железам разнообразных воздействий на организм, идущих из внешней среды. Характерные изменения в гормональной деятельности надпочечников, щитовидной железы и гонад, наступающие под воздействием не обычных раздражителей в так называемых реакциях "напряжения", или "стресса", многие исследователи также связывают с деятельностью ретикулярной формации.
Пути действия ретикулярной формации на периферические эндокринные железы изучены еще недостаточно. Имеющиеся экспериментальные данные пока не позволяют решить вопрос о том, оказывает ли она только общее активирующее действие на гипоталамус и переключает в него информацию от внешней среды и внутренних органов или же и сама принимает непосредственное участие в регуляции физиологической активности периферических эндокринных желез. Последнее предположение подтверждается отдельными наблюдениями. Известно, что после удаления гипофиза гормональная деятельность отдельных эндокринных желез полностью не прекращается, а сохраняется на уровне так называемой базальной активности, для которой характерна суточная ритмичность. Последняя, по-видимому, контролируется ретикулярной формацией. Рассмотренные факты приводят отдельных исследователей к заключению, что импульсы от ретикулярной формации могут достигать периферических желез без участия гипофиза. Таким образом, возможен и парагипо-физарный путь регуляции эндокринных желез. Ретикулярная формация не только оказывает влияние на гуморальную среду организма, но и сама реагирует на ее изменения. Это указывает на возможность участия ретикулярной формации в рассмотренном выше механизме обратных связей.
Реферат на тему:
Гипоталамус. Физиология гипоталамуса.
Выполнила: Андреева Юлия 4207
Гипоталамус
Гипоталамус - внешний подкорковый центр вегетативной нервной системы. Эта подбугорная область промежуточного мозга долгое время является важным объектом различных научных исследований.
В настоящее время для изучения различных структур мозга широко применяется метод вживления электродов. С помощью особой стереотаксической техники через трепанационное отверстие в черепе вводят электроды в любой заданный участок мозга. Электроды изолированны на всем протяжении, свободен только их кончик. Включая электроды в цепь, можно узколокально раздражать те или иные зоны.
В этой работе рассматриваются некоторые теоретические и физиологические аспекты данной области промежуточного мозга.
Общие функции гипоталамуса.
У позвоночных гипоталамус представляет собой главный нервный центр, отвечающий за регуляцию внутренней среды организма.
Филогенетически - это довольно старый отдел головного мозга, и поэтому у наземных млекопитающих строение его относительно одинаково, в отличие от организации таких более молодых структур, как новая кора и лимбическая система.
Гипоталамус управляет всеми основными гомеостатическими процессами. В то время как децеребрированному животному можно достаточно легко сохранить жизнь, для поддержания жизнедеятельности животного с удаленным гипоталамусом требуются особые интенсивные меры, так как у такого животного уничтожены основные гомеостатические механизмы.
Принцип гомеостаза заключается в том, что при самых разнообразных состояниях организма, связанных с его приспособлением к резко изменяющимся условиям окружающей среды (например, при тепловых или холодовых воздействиях, при интенсивной физической нагрузке и так далее), внутренняя среда остается постоянной и параметры ее колеблются лишь в очень узких пределах. Наличие и высокая эффективность механизмов гомеостаза у млекопитающих, и в частности у человека, обеспечивают возможность их жизнедеятельности при значительных изменениях окружающей среды. Животные, неспособные поддерживать некоторые параметры внутренней среды, вынуждены жить в более узком диапазоне параметров среды.
Например: Способность лягушек к терморегуляции настолько ограничена, что для того, чтобы выжить в условиях зимних холодов, им приходится опускаться на дно водоемов, где вода не замерзнет. Напротив, многие млекопитающие зимой могут вести столь же свободное существование, что и летом, несмотря на значительные колебания температуры.
Отсюда понятно - в связи со слабым развитием механизмов гомеостаза, эти животные менее свободны в своей жизнедеятельности, а если удален гипоталамус, следственно нарушены гомеостатические процессы, то для поддержания жизнедеятельности этого животного необходимы особые интенсивные меры.
Функциональная анатомия гипоталамуса.
Расположение гипоталамуса. Гипоталамус представляет собой небольшой отдел головного мозга весом около 5 грамм. Гипоталамус не обладает четкими границами, и поэтому его можно рассматривать как часть сети нейронов, протягивающейся от среднего мозга через гипоталамус к глубинным отделам переднего мозга, тесно связанным с филогенетически старой обонятельной системой. Гипоталамус является вентральным отделом промежуточного мозга, он лежит ниже (вентральнее) таламуса, образуя нижнюю половинку стенки третьего желудочка. Нижней границей гипоталамуса служит средний мозг, а верхней - конечная пластинка, передняя спайка и зрительный перекрест. Латеральнее гипоталамуса расположен зрительный тракт, внутренняя капсула и субталамические структуры.
Строение гипоталамуса. В поперечном направлении гипоталамус можно разделить на три зоны: 1) Перивентрикулярную; 2) Медиальную; 3) Латеральную.
Перивентрикулярная зона представляет собой тонкую полоску, прилежащую к третьему желудочку. В медиальной зоне различают несколько ядерных областей, расположенных в переднезаднем направлении. Преоптическая область филогенетически принадлежит к переднему мозгу, однако ее относят обычно к гипоталамусу.
От вентромедиальной области гипоталамуса начинается ножка гипофиза, соединяющаяся с адено- и нейрогипофизом. Передняя часть этой ножки носит название срединного возвышения. Там оканчиваются отростки многих нейронов преоптической и передней областей гипоталамуса, а также вентромедиального и инфундибулярного ядер (рис.1 - цифры: 1, 4, 5); здесь из этих отростков высвобождаются гормоны, поступающие через систему портальных сосудов к передней доле гипофиза. Совокупность ядерных зон, в которых содержатся подобные гормон-продуцирующие нейроны, носят название гипофизотропной области. (Рис.1 - участок, обозначенный прерывистой линией).
Отростки нейронов супраоптического и паравентрикулярного ядер (Рис. 1 - цифры 2 и 3) идут к задней доле гипофиза (эти нейроны регулируют образование и высвобождение окситоцина и АДТ, или вазопрессина). Связать конкретные функции гипоталамуса с его отдельными ядрами, за исключением супраоптического и паравентрикулярного ядер, невозможно.
В латеральном гипоталамусе не существует отдельных ядерных областей. Нейроны этой зоны диффузно располагаются вокруг медиального пучка переднего мозга, идущего в растрально-каудальном направлении от латеральных образований основания лимбической системы к передним центрам промежуточного мозга. Этот пучок состоит из длинных и коротких восходящих и нисходящих волокон.
Афферентные и эфферентные связи гипоталамуса. Организация афферентных и эфферентных связей гипоталамуса свидетельствует о том, что он служит важным интегративным центром для соматических, вегетативных и эндокринных функций.
Латеральный гипоталамус образует двухсторонние связи с верхними отделами ствола мозга, центральным серым веществом среднего мозга и с лимбической системой. Чувствительные сигналы от поверхности тела и внутренних органов поступают в гипоталамус по восходящим спинобульборетикулярным путям, которые ведут в гипоталамус, либо через таламус, либо через лимбическую область среднего мозга. Остальные афферентные сигналы поступают в гипоталамус по полисинаптическим путям, которые пока еще не все идентифицированы.
Эфферентные связи гипоталамуса с вегетативными и соматическими ядрами ствола мозга и спинного мозга образованы полиснаптическими путями, идущими в составе ретикулярной формации.
Медиальный гипоталамус обладает двусторонними связями с латеральным, и, кроме того, он непосредственно получает сигналы от некоторых остальных отделов головного мозга. В медиальной области гипоталамуса существуют особые нейроны, воспринимающие важнейшие параметры крови и спинномозговой жидкости: то есть эти нейроны следят за состоянием внутренней среды организма. Они могут воспринимать, например, температуру крови, водноэлектролитный состав плазмы или содержание гормонов в крови.
Через нервные механизмы медиальная область гипоталамуса управляет деятельностью нейрогипофиза, а через гормональные - аденогипофиза. Таким образом, эта область служит промежуточным звеном между нервной и эндокринной системой.
Гипоталамус и сердечно-сосудистая система.
При электрическом раздражении почти любого отдела гипоталамуса могут возникнуть реакции со стороны сердечно-сосудистой системы. Эти реакции, опосредованные в первую очередь симпатической системой, а также ветвями блуждающего нерва, идущими к сердцу, свидетельствуют о важном значении гипоталамуса для регуляции гемодинамики со стороны внешних нервных центров.
Раздражение какого-либо отдела гипоталамуса может сопровождаться противоположными изменениями кровотока в разных органах (например, увеличением кровотока в скелетных мышцах и одновременным снижением в сосудах кожи). С другой стороны, противоположные реакции сосудов какого-либо органа могут возникать при раздражении разных зон гипоталамуса. Биологическое значение подобных гемодинамических сдвигов можно понять лишь в том случае, если рассматривать их в связи с другими физиологическими реакциями, сопровождающими раздражение этих же подталомических зон. Иными словами, гемодинамические эффекты раздражения гипоталамуса входят в состав общих поведенческих или гомеостатических реакций, за которые отвечает этот центр.
В качестве примера можно привести пищевые и защитные поведенческие реакции, возникающие при электрическом раздражении ограниченных участков гипоталамуса. Во время защитного поведения артериального давления и кровоток в скелетных мышцах повышаются, а кровоток в сосудах кишечника снижается. При пищевом поведении возрастает артериальное давление и кровоток в кишечнике, а кровоток в скелетных мышцах уменьшается. Аналогичные изменения гемодинамических параметров наблюдаются и во время других реакций, возникающих в ответ на раздражение гипоталамуса, например при терморегуляторных реакциях или половом поведении.
За механизмы регуляции гемодинамики в целом (то есть артериального давления в большом кругу кровообращения, сердечного выброса и распределения крови), действующие по принципу следящих систем, отвечают нижние отделы ствола мозга. Эти отделы получают информацию от артериальных баро- и химорецепторов и механорецепторов предсердий и желудочков сердца и посылают сигналы к различным структурам сердечно-сосудистой системы по симпатическим и парасимпатическим эфферентным волокнам. Такая бульбарная саморегуляция гемодинамики в свою очередь управляется высшими отделами ствола мозга, и в особенности гипоталамуса. Эта регуляция осуществляется благодаря нервным связям между гипоталамусом и преганглионарными вегетативными нейронами. Высшая нервная регуляция сердечно-сосудистой системы со стороны гипоталамуса участвует во всех сложных вегетативных реакциях, для управления которыми простой саморегуляции недостаточно, к таким регуляциям можно отнести: терморегуляцию, регуляцию приема пищи, защитное поведение, физическую деятельность и так далее.
Приспособительные реакции сердечно-сосудистой системы во время работы. Механизмы приспособления гемодинамики при физической работе представляют теоретический и практический интерес. При физической нагрузке повышается сердечный выброс (главным образом в результате увеличения частоты сокращений сердца) и одновременно возрастает кровоток в скелетных мышцах. В то же время кровоток через кожу и органы брюшной полости снижается. Эти приспособительные циркуляторные реакции возникают практически одновременно с началом работы. Они осуществляются центральной нервной системой через гипоталамус.
У собаки при электрическом раздражении латеральной области гипоталамуса на уровне мамиллярных тел возникают точно такие же вегетативные реакции, как и при беге на тредбане. У животных в состоянии наркоза электрическое раздражение гипоталамуса может сопровождаться локомоторными актами и учащением дыхания. Путем небольших изменений положения раздражающего электрода можно добиться независящих друг от друга вегетативных и соматических реакций. Все эти эффекты устраняются при двусторонних поражениях соответствующих зон; у собак с такими поражениями исчезают приспособительные реакции сердечно-сосудистой системы к работе, и при беге на тредбане, такие животные быстро устают. Эти данные свидетельствуют о том, что в латеральной области гипоталамуса расположены группы нейронов, отвечающие за адаптацию гемодинамики к мышечной работе. В свою очередь эти отделы гипоталамуса контролируются корой головного мозга. Неизвестно, может ли осуществляться такая регуляция изолированным гипоталамусом, так как для этого необходимо, чтобы к гипоталамусу поступали особые сигналы скелетных мышц.
Гипоталамус и поведение.
Электрическое раздражение маленьких участков гипоталамуса сопровождается возникновением у животных типичных поведенческих реакций, которые столь же разнообразны, как и естественные видоспецифические типы поведения конкретного животного. Важнейшими из таких реакций являются оборонительное поведение и бегство, пищевое поведение (потребление пищи и воды), половое поведение и терморегуляторные реакции. Все эти поведенческие комплексы обеспечивают выживание особи и вида, и поэтому их можно назвать гомеостатическими процессами в широком смысле этого слова. В состав каждого из этих комплексов входят соматорный, вегетативный и гормональный компоненты.
При локальном электрическом раздражении каудального кольца у бодрствующей кошки возникает оборонительное поведение, которое проявляется в таких типичных соматорных реакциях, как выгибание спины, шипение, расхождение пальцев, выпускание когтей, а также вегетативными реакциями - учащенным дыханием, расширением зрачков и пилоэрекцией в области спины и хвоста. Артериальное давление и кровоток в скелетных мышцах при этом возрастают, а кровоток в кишечнике снижается. Такие вегетативные реакции связаны главным образом с возбуждением адренергических симпатических нейронов. В защитном поведении участвуют не только соматорная и вегетативная реакции, но и гормональные факторы.
При раздражении каудального отдела гипоталамуса болевые раздражения вызывают лишь фрагменты оборонительного поведения. Это свидетельствует о том, что нервные механизмы оборонительного поведения находятся в задней части гипоталамуса.
Пищевое поведение, также связанное со структурами гипоталамуса, по своим реакциям почти противоположно оборонительному поведению. Пищевое поведение возникает при местном электрическом раздражении зоны, расположенной на 2-3 мм дорзальнее зоны оборонительного поведения. В этом случае наблюдаются все реакции, характерные для животного в поисках пищи. Подойдя к миске, животное с искусственно вызванным пищевым поведением начинает есть, даже если оно не голодно, и при этом пережевывает даже несъедобные предметы.
При исследовании вегетативных реакций можно обнаружить, что такое поведение сопровождается увеличенным слюноотделением, повышением моторики и кровоснабжения кишечника и снижением мышечного кровотока. Все эти типичные изменения вегетативных функций при пищевом поведении служат как бы подготовительным этапом к приему пищи. Во время пищевого поведения повышается активность парасимпатических нервов желудочно-кишечного тракта.
Принципы организации гипоталамуса.
Данные систематических исследований гипоталамуса при помощи локального электрического раздражения свидетельствуют о том, что в этом центре существуют нервные структуры, управляющие самыми разнообразными поведенческими реакциями. В опытах с использованием других методов - например, разрушения или химического раздражения - это положение было подтверждено и расширено.
Пример: афагия (отказ от пищи), возникающая при поражениях латеральных областей гипоталамуса, электрическое раздражение которых приводит к пищевому поведению. Разрушение медиальных областей гипоталамуса, раздражение которых тормозит пищевое поведение (центров насыщения), сопровождается гиперфагией (чрезмерным потреблением пищи).
Области гипоталамуса, раздражение которых приводит к поведенческим реакциям, широко перекрываются. В связи с этим пока еще не удалось выделить функциональные или анатомические скопления нейронов, отвечающих за то или иное поведение. Так, ядра гипоталамуса, выявляемые при помощи нейрогистологических методов, лишь приблизительно соответствуют областям, раздражение которых сопровождается поведенческими реакциями. Таким образом, нервные образования, обеспечивающие формирование целостного поведения из отдельных реакций, не следует рассматривать как четко очерченные анатомические структуры (на что могло бы натолкнуть существование таких терминов, как “центр голода” и “центр насыщения”).
Нейронная организация гипоталамуса, благодаря которой это небольшое образование способно управлять множеством жизненно важных поведенческих реакций и нейрогуморальных регуляторных процессов, остается загадкой.
Возможно, группы нейронов гипоталамуса, отвечающие за выполнение какой-либо функции, отличаются друг от друга афферентными и эфферентными связями, медиаторами, расположением дендритов и тому подобное. Можно предположить, что в малоизученных нервных цепях гипоталамуса заложены многочисленные программы. Активизация этих программ под влиянием нервных сигналов от вышележащих отделов мозга (например, лимбической системы) и сигналов от рецепторов и внутренней среды организма может приводить к различным поведенческим и нейрогуморальным регуляторным реакциям.
Функциональные расстройства у людей с повреждениями гипоталамуса
У человека нарушения деятельности гипоталамуса бывают связаны главным образом с неопластическими (опухолевыми), травматическими или воспалительными поражениями. Подобные поражения могут быть весьма ограниченными, захватывая передний, промежуточный или задний отдел гипоталамуса. У таких больных наблюдаются сложные функциональные расстройства. Характер этих расстройств определяется, кроме всего прочего, остротой (например, при травмах), или длительностью (например, при медленно растущих опухолях) процесса. При ограниченных острых поражениях могут возникать значительные функциональные нарушения, в то время как при медленно растущих опухолях эти нарушения начинают проявляться лишь при далеко зашедшем процессе.
В таблице перечислены сложные функции гипоталамуса и нарушения этих функций. Расстройства восприятия, памяти и цикла сон/бодрствование частично связаны с повреждением восходящих и нисходящих путей, соединяющих гипоталамус с лимбической системой.
|
Передний отдел гипоталамуса и преоптическая область. |
Промежуточный отдел гипоталамуса. |
Задний отдел гипоталамуса. |
|
|
Регуляция цикла сон/бодрствование, терморегуляция, регуляция эндокринных функций. |
Восприятие сигналов, энергетический и водный баланс, регуляция эндокринных функций. |
Восприятие сигналов, поддержание сознания, терморегуляция, интеграция эндокринных функций. |
|
|
Поражения: а) Острые |
Бессоница, гипертермия, несахарный диабет. |
Гипертермия, несахарный диабет, эндокринные нарушения. |
Сонливость, эмоциональные и вегетативные нарушения, пойкилотермия. |
|
б) Хронические |
Бессоница, сложные эндокринные расстройства (например раннее половое созревание), эндокринные расстройства, связанные с поражением срединного возвышения, гипотермия, отсутствие чувства жажды. |
Медиальный: нарушения памяти, эмоциональные расстройства, гиперфагия, ожирение, эндокринные нарушения. Латеральный: эмоциональные нарушения, потеря аппетита, истощение, отсутствие чувства жажды. |
Амнезия, эмоциональные нарушения, вегетативные расстройства, сложные эндокринные нарушения (раннее половое созревание). |
Список использованной литературы.
Физиология человека. Том 1, под ред.акад. П.Г.Костюка. “Мир”, 1985.
Воробьева Г.А., Губарь Л.В., Сафьянникова С.Б., Анатомия и физиология.
Ермолаев И.И., Возрастная физиология.
Фомин А.Б., Физиология человека, “Просвещение”,1995.
Гипоталамус (hypothalamus) - отдел промежуточного мозга, которому принадлежит ведущая роль в регуляции многих функций организма, и прежде всего постоянства внутренней среды, гипоталамус является высшим вегетативным центром, осуществляющим сложную интеграцию функций различных внутренних систем и их приспособление к целостной деятельности организма, играет существенную роль в поддержании оптимального уровня обмена веществ и энергии, в терморегуляции, в регуляции деятельности пищеварительной, сердечно-сосудистой, выделительной, дыхательной и эндокринной систем. Под контролем гипоталамуса находятся такие железы внутренней секреции, как гипофиз , щитовидная железа , половые железы (см. Яичко , Яичники ), поджелудочная железа , надпочечники и др.
Гипоталамус расположен книзу от таламуса под гипоталамической бороздой. Его передней границей являются зрительный перекрест (chiasma opticum), терминальная пластинка (lamina terminalis) и передняя спайка (commissura ant.). Задняя граница проходит позади нижнего края сосцевидных тел (corpora mamillaria). Кпереди клеточные группы гипоталамуса без перерыва переходят в клеточные группы пластинки прозрачной перегородки (lamina septi pellucidi).
Проводящие пути тесно связывают гипоталамус с соседними структурами головного мозга . Кровоснабжение ядер гипоталамуса осуществляется веточками артериального круга головного мозга. Взаимосвязь между гипоталамусом и аденогипофизом происходит через портальные сосуды аденогипофиза. Характерной особенностью кровеносных сосудов гипоталамуса является проницаемость их стенок для крупных молекул белков.
Несмотря на небольшие размеры гипоталамуса , его строение отличается значительной сложностью Группы клеток образуют отдельные ядра гипоталамуса (см. илл. К ст. Головной мозг ). У человека и других млекопитающих в гипоталамусе обычно различают 32 пары ядер. Между соседними ядрами существуют промежуточные нервные клетки или их небольшие группы, поэтому физиологическое значение могут иметь не только ядра, но и некоторые межъядерные гипоталамические зоны. Ядра гипоталамуса образуются нервными клетками, не обладающими секреторной функцией, и нейросекреторными клетками. Нейросекреторные нервные клетки сконцентрированы непосредственно около стенок III желудочка мозга. По своим структурным особенностям эти клетки напоминают клетки ретикулярной формации и продуцируют физиологически активные вещества - гипоталамические нейрогормоны .
В гипоталамусе выделяют три нерезко разграниченные области: переднюю, среднюю и заднюю. В передней области гипоталамуса сосредоточены нейросекреторные клетки, где они образуют с каждой стороны надзрительное (nucl. supraopticus) и паравентрикулярное (nucl. paraventricularis) ядра. Надзрительное ядро состоит из клеток, лежащих между стенкой III желудочка мозга и дорсальной поверхностью зрительного перекреста. Паравентрикулярное ядро имеет вид пластинки между сводом (fornix) и стенкой III желудочка мозга. Аксоны нейронов паравентрикулярного и надзрительного ядер, образуя гипоталамо-гипофизарный пучок, достигают задней доли гипофиза, где накапливаются гипоталамические нейрогормоны, оттуда они поступают в кровоток.
Между надзрительным и паравентрикулярным ядрами расположены многочисленные одиночные нейросекреторные клетки или их группы. Нейросекреторные клетки надзрительного ядра гипоталамуса вырабатывают преимущественно антидиуретический гормон (вазопрессин), а паравентрикулярного ядра - окситоцин.
В средней области гипоталамуса , вокруг нижнего края III желудочка мозга, лежат серобугорные ядра (nucll. tuberaies), дуговидно охватывающие воронку (infundibulum) гипофиза. Кверху и немного латеральнее от них находятся крупные вентромедиальные и дорсомедиальные ядра.
В задней области гипоталамуса расположены ядра, состоящие из рассеянных крупных клеток, среди которых находятся скопления мелких клеток К этому отделу относятся также медиальные и латеральные ядра сосцевидного тела (nucll. corporis mamillaris mediales et laterales), которые на нижней поверхности промежуточного мозга имеют вид парных полушарий. Клетки этих ядер дают начало одной из так называемых проекционных систем гипоталамуса в продолговатый и спинной мозг. Наиболее крупным клеточным скоплением является медиальное ядро сосцевидного тела. Кпереди от сосцевидных тел выступает дно III желудочка мозга в виде серого бугра (tuber cinereum), образованного тонкой пластинкой серого вещества. Этот выступ вытягивается в воронку, переходящую в дистальном направлении в гипофизарную ножку и далее в заднюю долю гипофиза. Расширенная верхняя часть воронки - срединное возвышение - выстлано эпендимой, за которой идут слой нервных волокон гипоталамо-гипофизарного пучка и более тонкие волокна, берущие начало от ядер серого бугра. Наружная часть срединного возвышения образована опорными нейроглиальными (эпендимными) волокнами, между которыми залегают многочисленные нервные волокна. В этих нервных волокнах и около них наблюдается отложение нейросекреторных гранул. Т.о., гипоталамус образован комплексом нервно-проводниковых и нейросекреторных клеток. В связи с этим регулирующие влияния гипоталамусу передаются к эффекторам, в т.ч. и к железам внутренней секреции, не только с помощью гипоталамических нейрогормонов, переносимых с током крови и, следовательно, действующих гуморально, но и по эфферентным нервным волокнам.
Значительна роль гипоталамуса в регуляции и координации функций вегетативной нервной системы. В регуляции функции ее симпатической части участвуют ядра задней области гипоталамуса , а функции парасимпатической части вегетативной нервной системы регулируют ядра его передней и средней областей. Стимуляция передней и средней областей гипоталамуса вызывает реакции, характерные для парасимпатической нервной системы - урежение сердцебиений, усиление перистальтики кишечника, повышение тонуса мочевого пузыря и др., а раздражение задней области гипоталамуса проявляется усилением симпатических реакций - учащением сердцебиений и т.д.
С состоянием вегетативной нервной системы тесно связаны вазомоторные реакции гипоталамического происхождения. Различные виды артериальной гипертензии, развивающиеся после стимуляции гипоталамуса , обусловлены комбинированным влиянием симпатической части вегетативной нервной системы и выделением адреналина надпочечниками , хотя в данном случае нельзя исключить влияние нейрогипофиза, особенно в генезе устойчивой артериальной гипертензии.
С физиологической точки зрения гипоталамус имеет ряд особенностей, прежде всего это касается его участия в формировании поведенческих реакций, важных для сохранения постоянства внутренней среды организма (см. Гомеостаз ). Раздражение гипоталамуса приводит к формированию целенаправленного поведения - пищевого, питьевого, полового, агрессивного и т.п. Гипоталамусу принадлежит главная роль в формировании основных влечений организма (см. Мотивации ). В некоторых случаях при повреждении верхнемедиального ядра и серобугровой области гипоталамуса наблюдают чрезмерное ожирение как результат полифагии (булимий) или кахексию. Повреждение задних отделов гипоталамуса вызывает гипергликемию. Установлена роль надзрительного и паравентрикулярного ядер в механизме возникновения несахарного диабета (см. Диабет несахарный ). Активация нейронов латерального гипоталамуса вызывает формирование пищевой мотивации. При двустороннем разрушении этого отдела пищевая мотивация полностью устраняется.
Обширные связи гипоталамуса с другими структурами головного мозга способствуют генерализации возбуждений, возникающих в его клетках. Гипоталамус находится в непрерывных взаимодействиях с другими отделами подкорки и корой головного мозга. Именно это лежит в основе участия гипоталамуса в эмоциональной деятельности (см. Эмоции ). Кора головного мозга может оказывать тормозящий эффект на функции гипоталамуса . Приобретенные корковые механизмы подавляют многие эмоции и первичные побуждения, формирующиеся с его участием. Поэтому декортикация нередко приводит к развитию реакции «мнимой ярости» (расширение зрачков, тахикардия, развитие внутричерепной гипертензии, усиление саливации и т.д.).
Гипоталамус является одной из главных структур, участвующих в регуляции смены сна и бодрствования. Клиническими исследованиями установлено, что симптом летаргического сна при эпидемическом энцефалите обусловлен именно повреждением гипоталамуса . В поддержании состояния бодрствования решающую роль играет задняя область гипоталамус а . Обширное разрушение средней области гипоталамуса в эксперименте приводило к развитию длительного сна. Нарушение сна в виде нарколепсии объясняется поражением гипоталамуса и ростральной части ретикулярной формации среднего мозга.
Гипоталамус играет важную роль в терморегуляции . Разрушение задних отделов гипоталамуса приводит к стойкому снижению температуры тела.
Клетки гипоталамуса обладают способностью трансформировать гуморальные изменения внутренней среды организма в нервный процесс. Центры гипоталамуса характеризуются выраженной избирательностью возбуждения в зависимости от различных изменений состава крови и кислотно-щелочного состояния, а также нервных импульсов из соответствующих органов. Возбуждение в нейронах гипоталамуса , обладающих избирательной рецепцией по отношению к константам крови, возникает не сразу, как только изменится какая-либо из них, а через определенный промежуток времени. Если же изменение константы крови поддерживается длительно, то в этом случае возбудимость нейронов гипоталамуса быстро поднимается до критической величины и состояние этого возбуждения поддерживается на высоком уровне все время, пока существует изменение константы. Возбуждение одних клеток гипоталамуса может возникать периодически через несколько часов, как, например, при гипогликемии, других - через несколько суток или даже месяцев, как, например, при изменении содержания в крови половых гормонов.
Информативными методами исследования гипоталамуса являются плетизмографические, биохимические, рентгенологические исследования и др. Плетизмографические исследования (см. Плетизмография ) выявляют широкий спектр изменений в гипоталамуса - от состояния вегетативной сосудистой неустойчивости и парадоксальной реакции до полной арефлексии. При биохимических исследованиях у больных с поражением гипоталамуса независимо от его причины (опухоль, воспалительный процесс и др.) часто определяется увеличение содержания катехоламинов и гистамина в крови, увеличивается относительное содержание a -глобулинов и снижается относительное содержание b -глобулинов в сыворотке крови, изменяется экскреция с мочой 17-кетостероидов. При различных формах поражения гипоталамус а проявляются нарушения терморегуляции и интенсивности потоотделения. Поражение ядер гипоталамуса (преимущественно надзрительного и паравентрикулярного) наиболее вероятно при заболеваниях желез внутренней секреции, черепно-мозговых травмах, приводящих к перераспределению цереброспинальной жидкости, опухолях, нейроинфекциях, интоксикациях и др. Вследствие повышения проницаемости стенок сосудов при инфекциях и интоксикациях гипоталамические ядра могут подвергаться патогенным воздействиям бактериальных и вирусных токсинов и химических веществ, циркулирующих в крови. Особенно опасны в этом отношении нейровирусные инфекции. Поражения гипоталамуса наблюдаются при базальном туберкулезном менингите, сифилисе, саркоидозе, лимфогранулематозе, лейкозах.
Из опухолей гипоталамуса наиболее часто встречаются различного вида глиомы, краниофарингиомы, эктопические пинеаломы и тератомы, менингиомы: в гипоталамусе прорастают супраселлярные аденомы гипофиза . Клинические проявления и лечение нарушений функций и заболеваний гипоталамуса - см. Гипоталамо-гипофизарная недостаточность , Гипоталамические синдромы , Адипозогенитальная дистрофия , Иценко - Кушинга болезнь , Диабет несахарный , Гипогонадизм , Гипотиреоз и др.
Библиогр.: Бабичев В.Н. Нейроэндокринология пола. М., 1981; он же, Нейрогормональная регуляция овариального цикла, М., 1984; Шрейбер В. Патофизиология желез внутренней секреции, пер. с чешск., Прага, 1987.
Эндокринная система состоит из группы желез внутренней секреции. Деятельностью этих желез управляют две железы - гипоталамус и гипофиз . Они вырабатывают и выбрасывают в кровь гормоны - химические вещества, влияющие на метаболизм, развитие организма и рост, а также деятельность различных органов и тканей.
. Является отделом головного мозга , объединяющим нервную систему с эндокринной, отдел регулируюет работу гипофиза, а через него происходит и контроль работы всей эндокринной системы.
Занимается выработкой гормонов, оказывающих прямое воздействие на ткани организма. Также именно гипофиз контролируют работу других желез эндокринной системы.

Меланоцитстимулирующий гормон;(МСГ)
Область действия: Кожа.
Функции: Стимулирует выработку меланоцитов, влияющих на цвет кожи.
Антидиуретический гормон, или вазопрессин (АДГ)
Область действия: Почки.
Функции: Удерживает воду в почках, регулирует артериальное давление.
Гормон роста, или соматотропин (ГР, СТГ или РГ);
Область действия: Весь организм.
Функции: Стимулирует рост мышц, костей и органов в детстве и во время полового созревания.
Тиреотропин (ТТГ)
Область действия: Щитовидная железа.
Функции: Стимулирует деятельность щитовидной железы.
Окситоцин
Область действия: Матка
Функции: Провоцирует сокращения матки во время родов.
Адренокортикотропин (АКТГ)
Область действия: Надпочечники.
Функции: Стимулирует выработку кортикостероидов надпочечными железами.
Пролактин (ЛТГ)
Область действия: Грудь.
Функции: Провоцирует выработку молока грудными железами после родов.
Гонадотропины
;Фолликулостимулирующий гормон (ФСГ)
;Лютеинизирующий гормон, или гормон, стимулирующий интерстнциальные клетки (ЛГ или ГСИК)
Область действия: Гонады (яйцеклетки и яички).
Функции: Регулирует созревание сперматозоидов и яйцеклеток, а также выработку половых гормонов.